Ключевые слова: крысы, парадоксальный сон, циркуляторная ишемия
Исследование проведено на нелинейных крысах, переживших окклюзию одной или двух общих сонных артерий (N=30, около 30% от оперированных). Контролем служили животные, оперированные без окклюзии сонных артерий (N=32).
Показано значительное увеличение суммарной продолжительности сна за 3 часа у ишемизированных животных по сравнению с контролем, за счет значительного (в 8-9 раз) увеличения длительности парадоксальной стадии сна (ПС). В группе ишемизированных животных в первый послеоперационный день наблюдалось резкое увеличение длительности ПС и затем постепенное уменьшение ПС в период с первого по сороковой послеоперационные дни. Полученные результаты указывают на участие ПС в реализации собственных репаративных функций головного мозга.
INCREASE IN PARADOXICAL SLEEP TIME AFTER CAROTID OCCLUSION IN RATS
V.V Loginov., V.B. Dorokhov
Institute of Immunopatology;
Institute of Higher Nervous Activity and Neurophysiology. Russian Academy of Science. Moscow.
e - mail: vikloginov@yandex.ru, vbdorokhov@mail.ru
Key words: paradoxical sleep, experimental ischemia, rat
The research is carried out on nonlinear rat survived occlusion of one or two carotid artery (N=30, about 30% survived). Control group: animals (N=32) without occlusion of carotid artery.
It is shown substantial growth of total sleep duration during 3 hours at animals with ischemia in comparison with the control groups. It occurred at the expense of substantial growth (in 8-9 times) of paradoxical phase of sleep (PS). Animals with ischemia in first day after surgery was observed the sharp increase of PS duration and then gradual reduction of PS duration from first on fortieth days. In our opinion the received results about significant lengthening of the PS duration at an animal successfully survived experimental ischemia to displays participation PS in realization of mechanisms of own brain reparation.
В конце 70-х годов И. Освальдом была сформулирована гипотеза, согласно которой в период естественного сна происходит восстановление "резервного" потенциала клеток. Исследуя изменения структуры сна, он показал, что в период медленного сна (МС) реализуются анаболические процессы всего организма, включая головной мозг, а в период парадоксального сна (ПС) - синтетические процессы в ЦНС [4]. Это означает, что у переживающих циркуляторную ишемию крыс [3,6], в клетках мозга которых нарушается энергетический метаболизм, следует ожидать объективные количественные изменения структуры естественного сна, связанные с активацией собственных восстановительных функций мозга. Для проверки этого предположения изучали структуру естественного сна у крыс, успешно переживших циркуляторную ишемию в восстановительном периоде после окклюзии сонных артерий. Следует отметить особенность системы кровообращения головного мозга лабораторных белых крыс, которая заключается в том, что сосудистая система виллизиева круга у этих животных слабо развита и обеспечивает кровью, в основном, переднюю часть головного мозга [2].
Исследование проведено на нелинейных крысах-самцах массой 180-200 г., переживших блокирование кровотока в виллизиевом круге (около 30% от общего числа оперированных животных). Блокировку кровотока осуществляли перевязкой одной или обеих общих сонных артерий под наркозом (нембутал 40 мг/кг), что является распространенной экспериментальной моделью церебральной ишемии. Электроды вживляли: в передний и задний отделы коры, в гиппокамп; для записи электромиограммы - в шейную мышцу. Индифферентным электродом служил металлический винт - закрепленный в носовой кости. Регистрацию стадий и фаз сна осуществляли в условиях, не ограничивающих двигательную активность животных, с 9 до 12 часов. В трехчасовой записи визуально оценивали стадии медленного сна и фазу ПС. В фазе медленного сна отмечали поверхностную (с преобладанием веретен) и глубокую стадию (с преобладанием дельта-волн). ПС определяли по моноритмичному (слабо модулированному по частоте) гиппокампальному тета-ритму, максимальному снижению в это время мышечной активности и быстрым движениям глаз, отражающимся в переднем кортикальном отведении ЭЭГ.
Исследовали две группы ишемизированных животных: 1) с односторонней перевязкой сонной артерии (оперированных N=47, выживших N=14) и 2) с двусторонней перевязкой сонных артерий (оперированных N=58, выживших N=16). Результаты анализировались только у животных проживших более 40 дней. В качестве двух контрольных групп использовали: 1) животных оперированных только для вживления регистрирующих электродов (N=15), и 2) животных, у которых операция вживления электродов сочеталось с начальным этапом операции окклюзии сонных артерий, и ограничивалась только этапом доступа к сонным артериям (N=17).
Сравнение продолжительности МС и ПС у двух исследуемых групп между собой не выявило существенных различий. Значимых различий не было обнаружено и при сравнении двух контрольных групп между собой. Достоверные различия были обнаружены только при сравнении каждой из исследуемых групп с контролем 1 и контролем 2 (таблица 1).
Общим для всех стадий и фаз сна было увеличение их продолжительности так, что в целом суммарная продолжительность сна за 3 часа у ишемизированных животных была существенно больше в обеих исследуемых группах в сравнении с каждым из контролей (уровень значимости различий - при р<0,001). Однако вклад анализируемых показателей в суммарную продолжительность сна оказался неодинаковым. Увеличение показателей медленного сна было на уровне тенденции. Существенное же увеличение суммарной продолжительности сна произошло за счет 8-9-кратного увеличения ПС в первый послеоперационный день (р<0,001 во всех сравнениях, см. таблицу).
На рисунке 1 представлена динамика продолжительности ПС у крыс после операции. Видно, что в восстановительном периоде в контрольных группах на пятый послеоперационный день значения продолжительности ПС практически стабилизируются (нормализуются). В отличие от этого, ПС в группе ишемизированных животных был существенно более выражен в остром периоде и значительно медленнее претерпевал обратную динамику после операции.
Полученные результаты подтверждают гипотезу И.Освальда [4] об отражении в ПС активности восстановительных процессов головного мозга. Прежде всего, это следует из того, что исследовались животные, успешно пережившие экспериментальную ишемию, т.е. был произведен своеобразный "направленный отбор" лабораторных крыс с наиболее эффективно работающими репаративными механизмами головного мозга. При этом в восстановительном периоде не применялось никакой фармакотерапии и, следовательно, нормализацию ПС в это время следует отнести за счет собственных восстановительных процессов мозга. Это согласуется с результатами о взаимосвязи индивидуальных типологических особенностях поведения животных с определенной спецификой энергетического метаболизма мозга [3,6].
Проведенный нами анализ опубликованных данных о возможных причинах увеличения продолжительности ПС [7-9, 11] показал, что в большинстве случаев эффект может быть связан с переходом деятельности мозга в режим преобладания работы филогенетически более древних механизмов, что хорошо согласуется с представлениями о филогенетически древних механизмах происхождения парадоксального сна, развиваемых В.М.Ковальзоном [10]. По-видимому, в обычных условиях для нормализации работы мозга достаточно непродолжительного эпизодического включения механизмов парадоксального сна. В случае же такого экстремального воздействия, как ишемия головного мозга, происходит переход организма в режим преобладания функционирования филогенетически более древних механизмов и репаративная функция парадоксального сна включается на более продолжительное время, что позволяет, как было показано, избежать летального исхода у части животных.
Полученные нами экспериментальные данные о существенной роли ПС для восстановления центральных функций подтверждается и клиническими наблюдениями, опубликованными в литературе. Известно, что полисомнографический мониторинг играет важную роль в определении витального прогноза при мозговом инсульте. В частности, выраженная сегментации сна при этом заболевании, частые переходы к бодрствованию из разных стадий и фаз, а также отсутствие ПС и/или дельта-сна, являются прогностически неблагоприятными признаками. Такая закономерность выявлена не только для острой фазы инсультов (1-я неделя), но и для более отдаленного периода (3-4 недели). При этом перечисленные прогностически неблагоприятные признаки наблюдаются у больных с локализацией поражения в срединных или стволовых структурах головного мозга [1].
Таким образом, сопоставление полученного результата об увеличение продолжительности парадоксальной фазы сна при экспериментальной ишемии мозга с клиническими наблюдениями у больных с сосудистыми нарушениями мозга [1] свидетельствует о возможности использования полученного феномена в практике лечения больных инсультами. Представляются перспективными следующие направления использования полученного результата: во-первых - в качестве неинвазивного метода функциональной оценки ишемического очага (его динамики в процессе лечения, эффективности терапии и прогноза исхода), во-вторых - для оценки влияния действия препаратов различной направленности на собственные восстановительные процессы мозга больных инсультом.
Список литературы:
- 1. Гасанов Р.Л., Гитлевич Т.Р., Левин Я.И., Лесняк В.Н. Динамика структуры ночного сна у больных мозговым инсультом // конф. "Актуальные проблемы сомнологии". Тезисы докладов, Москва, 1998, C. 29.
- 2. Гусев Е.И., Боголепов Н.Н., Бурд Г.С., Румянцева С.А., Ультраструктурные аспекты острой церебральной ишемии при воздействии барбитуратов // Ж. Невропатологии и Психиатрии, 1982, Т. 82, № 9, С. 8-14.
- 3. Косицын Н.С., Ионкина Е.Г., Сердюченко В.М. Реакция апикальных дендритов пирамидных нейронов коры мозга на экспериментальную ишемию. Докл. АН СССР, 1988, Т. 298, № 1, С. 247-250.
- 4. Освальд И. Анаболическая функция сна человека. // "Саморегуляция процесса сна", Л. Наука, 1977, С. 12-14.
- 5. Полежаев Л.В., Александрова М.А., Аллотрансплантация эмбриональной ткани мозга в головной мозг взрослых млекопитающих при гипоксической гипоксии. // Ж. Невропатологии и психиатрии, 1983, т. 83, № 7, С. 984-997.
- 6. Саркисова К.Ю., Куликов М.А., Ноздрачева Л.В., Коломейцева И.А. Значение индивидуально-типологических особенностей поведения и энергетического метаболизма мозга для прогнозирования течения экспериментальной патологии (на примере циркуляторной гипоксии мозга). // Нарушения высшей нервной деятельности, их патогенез и нейропептидная коррекция. М. Наука. 1992. С. 113-127.
- 7. Bell-Pedersen D., Shinohara M.L., Loros J.J. Circadian clock-controlled genes isolated from Neurospora crassa are late night-to early morning-specific // 1996. V. 93. P. 13096-13101.
- 8. Cameron H.A., McKay R. Stem cells and neurogenesis in the adult brain // Curr. Opin. Neurobiol. -1998. V. 8. P. 677-680.
- 9. Dunlop J.C. Chronic hypertension and perinatal mortality. // Annu. Rev. Genet. 1996. V. 30. P. 570 -601
- 10. Kovalzon V.M. Origin of sleep. // Neurobiology of Sleep-Wakefulness Cycle. 2002. V.2. N1. P.33-36 (http://nswc.gateway.ge).
- 11. Vescovi A.L., Snyder E.Y. Establishment and properties of neural stem cell clones: plasticity in vitro and in vivo. // Brain Pathol. 1999.V. 9. P. 569-598.
Таблица 1.
Изменение продолжительности фазы парадоксального сна и медленного сна у крыс после перевязки одной и двух общих сонных артерий в первый день после операции. Продолжительность фаз сна указана в процентах от суммарной продолжительности сна за 3 часа.
| Группы | Кол-во | Фаза МС | Фаза ПС |
| (N) | Поверхност-ная
стадия | Глубокая
стадия | |
| Контроль I | 15 | 24,0+1,4 | 8,3+1,3 | 2,2+0,4 |
| Контроль II | 17 | 25,2+1,9 | 9,1+1,7 | 2,4+0,5 |
Окклюзия одной
артерии | 14 | 28,9+3,1 | 12,1+2,8 | 16,9+2,0* |
Окклюзия обеих
артерий | 16 | 31,2+3,8 | 10,7+3,7 | 18,0+1,8 * |
*- значимость отличий от контроля 1 и от контроля 2 на уровне р<0,001 (t- критерий Стьюдента)
Рис. 1.
Динамика продолжительности фазы парадоксального сна у крыс, успешно переживающих перевязку одной и обеих общих сонных артерий.
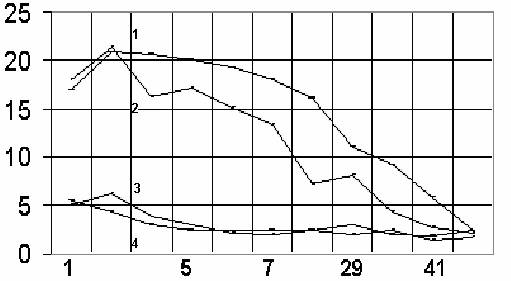
Первая (1) и вторая (2) линии сверху - перевязка одной и двух общих сонных артерий соответственно. Третья (3) и четвертая (4) линии - животные после стандартной операции вживления электродов и ложно оперированные животные. Ордината - продолжительность парадоксального сна в % от суммарной продолжительности сна за 3 часа. Абсцисса - номер послеоперационного дня.